|
Roy Larick Shore Cultural Centre, Euclid, Ohio
Russell L. Ciochon Dept. of Anthropology, University of Iowa,
Iowa City, Iowa
Yahdi Zaim Dept of Geology, Institute of Technology,
Bandung, Indonesia
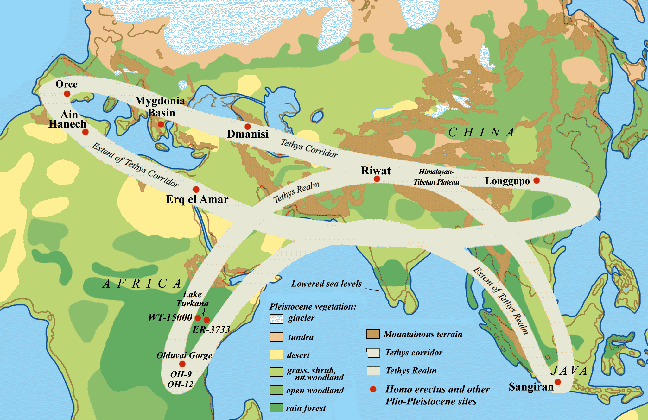 Fig.1: The Tethys Corridor, shown with generalized Pleistocene vegetation
zones and Homo erectus sites. Fig.1: The Tethys Corridor, shown with generalized Pleistocene vegetation
zones and Homo erectus sites.
Fossils of
“Java Man” were first found on that Southeast Asian island 112
years ago. Although their origin was not recognized at the time, the finds
represented Homo erectus, an early human species that presumably evolved
in equatorial Africa more than 2 million years ago. But the how, why, and
when of Homo erectus’ trek from Africa to island Southeast Asia
have always been difficult questions.
The trek was indeed complex, and it appears that Homo erectus’
arrival to current Java is an important factor for understanding why the
species left highland tropical Africa. We hope to shed light on the larger
issues by delving into Homo erectus’ significant relationship
with volcanic and otherwise highly unstable landscapes in far Southeast Asia.
Homo erectus seems to have sought out unstable landscapes, and this
preference may explain this hominid’s departure from Africa before 1.8
million years ago (mya) and its arrival to extreme Southeast Asia not long
thereafter.
Our theory
is based on two premises. First is the fact that early Homo erectus
fossils are always found in the context of volatile geology and ragged
geography. Second, the East African Rift and extreme Southeast Asia are endpoints
o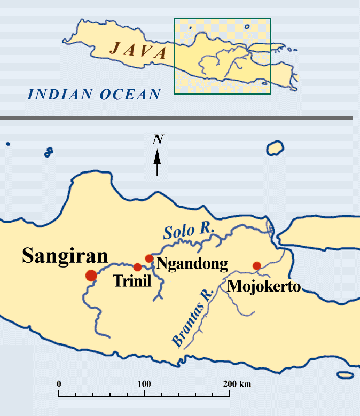 n a grand east-west geotectonic pathway called the Tethys corridor (fig.1).
During a rather brief period called the Olduvai subchron (1.98-1.79 mya),
the Tethys corridor was extremely unstable. Homo erectus and companion
mammals took advantage of open linear landscapes to migrate north from the
Rift to the Caucasus, and then both ways across the Tethys corridor - west
toward Gibraltar, east to the Himalayan fore slope, and then far east to
current Java. By the end of the Olduvai subchron, Homo erectus had
dispersed throughout the greater Tethys realm. n a grand east-west geotectonic pathway called the Tethys corridor (fig.1).
During a rather brief period called the Olduvai subchron (1.98-1.79 mya),
the Tethys corridor was extremely unstable. Homo erectus and companion
mammals took advantage of open linear landscapes to migrate north from the
Rift to the Caucasus, and then both ways across the Tethys corridor - west
toward Gibraltar, east to the Himalayan fore slope, and then far east to
current Java. By the end of the Olduvai subchron, Homo erectus had
dispersed throughout the greater Tethys realm.
Fig.2:
Map showing concentration of sites with Homo erectus fossil
finds in Central and East Java.
The immediate “how” of the Southeast Asian trek is perhaps the
easiest to answer. We now know that Homo erectus did not navigate to present
Java, but rather walked the length of the emergent Sunda continental shelf
off East Asia’s present south coast. Currently, the Indonesian
archipelago’s 14,000 tropical islands constitute the emergent landmass
(fig.3), but in the last 2 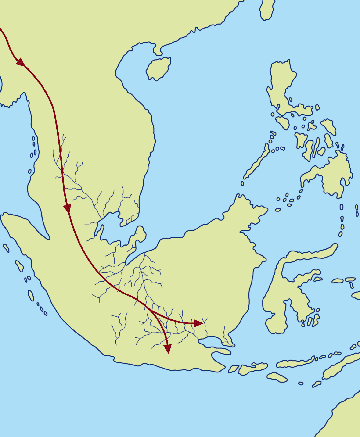 million years, the Sunda subcontinent (fig.4)
has sometimes included much of million years, the Sunda subcontinent (fig.4)
has sometimes included much of 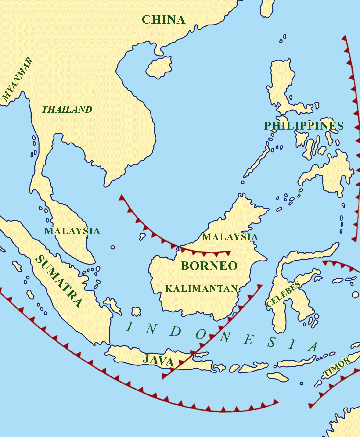 the intervening sea bottom. Arriving to the
area of present-day Thailand, Homo erectus groups spread south along
a broad valley between the current Malay Peninsula and Sumatra. And they
continued on to current Central Java, the extreme south coast of Sunda (fig.2). the intervening sea bottom. Arriving to the
area of present-day Thailand, Homo erectus groups spread south along
a broad valley between the current Malay Peninsula and Sumatra. And they
continued on to current Central Java, the extreme south coast of Sunda (fig.2).
Fig.3 (left): Southeast Asia today with subduction zones (after
Djubiantono and Sémah 1993). Fig.4 (right): Schematic map of “Sundaland” at 2-1.5 mya,
with probable migration path of Homo erectus (after Djubiantono and
Sémah 1993).
 Much of Central
Java lies directly above Sunda’s highly volcanic subduction zone. Active
volcanoes form the spine of the entire island but are especially large in
the central zone (fig.5), and their ash is slightly alkaline. Central Java
is consequently blessed with rich volcanic soils; they are young and
“sweet.” Using this resource of the central district, the entire island has supported a much larger historical human population than Sumatra,
the Malay Peninsula, or Borneo - Java is earth’s most densely inhabited
agricultural island. Central Java’s soil wealth probably explains why
Homo erectus may have explored much of Sunda, but gravitated to this
coast. As a hunter, this hominid also took advantage of eruptions that randomly
cleared patches of rainforest. Such events provided rich graze for the large
mammals upon which the early humans preyed. Much of Central
Java lies directly above Sunda’s highly volcanic subduction zone. Active
volcanoes form the spine of the entire island but are especially large in
the central zone (fig.5), and their ash is slightly alkaline. Central Java
is consequently blessed with rich volcanic soils; they are young and
“sweet.” Using this resource of the central district, the entire island has supported a much larger historical human population than Sumatra,
the Malay Peninsula, or Borneo - Java is earth’s most densely inhabited
agricultural island. Central Java’s soil wealth probably explains why
Homo erectus may have explored much of Sunda, but gravitated to this
coast. As a hunter, this hominid also took advantage of eruptions that randomly
cleared patches of rainforest. Such events provided rich graze for the large
mammals upon which the early humans preyed.
Fig.5: The Solo Basin, a segment of Java’s east-west trending
Central Depression, the current magmatic zone of the Indonesian volcanic
island arc (after Larick et al. 2001).
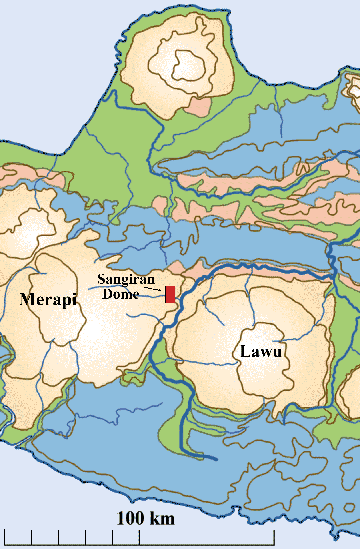
The Sangiran Dome: Central Java’s Sangiran dome lies squarely
astride the subduction zone; large volcanoes rise to the east (Lawu) and
to the west (Merapi) (figs.5,6). The dome is a window into one of earth’s
more important early human fossil beds and is now a UNESCO World Heritage
Site. During the last century, the dome’s heavily eroded cliff faces
have yielded more than 80 fragments of Homo erectus skeletons (Larick,
Ciochon, and Zaim 1999). The fossils represent an early human occupation
of about 500,000 years’ duration as Sunda emerged from the Java Sea.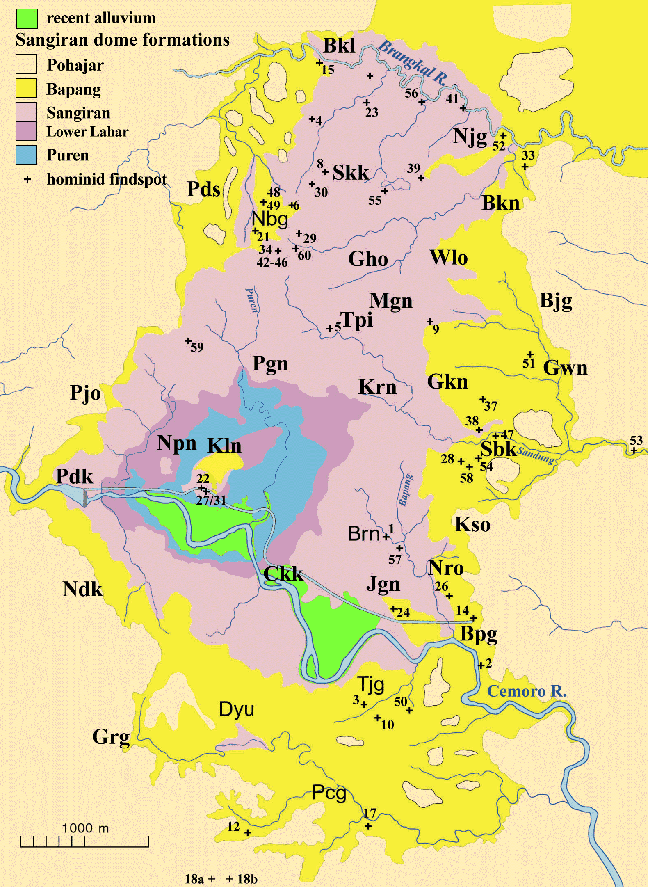
Two million
years ago, the area of the current dome lay on the southern edge of the submerged
Sunda shelf. The edge emerged from the Java Sea as three base geological
factors began to interplay. One was the slow but constant folding of Asian
crust as the Indian Oceanic plate slid or subducted under it at the Java
trench just south of the island. Another was the buildup of debris from volcanoes
venting above the subduction zone. The critical factor was the coming of
Pleistocene “Ice Age” around 2 mya. At this point, glacial ice
accumulated rapidly on the northern hemisphere’s continental masses.
So much water was pulled out of the earth’s atmosphere and locked up
as ice that sea levels receded across the globe. While sea levels have fluctuated
with glacial cycles over this period, the net effect has been lower water
and a more emergent Sunda shelf.
Fig.6: Sangiran Dome, showing sedimentary levels and documented hominid
fossil findspots. Abbreviations mark village localities (after Larick
et al. 2001).
The lowest formation exposed in the dome, the Puren limestone, represents
this earliest stage of emergence. In its upper reaches, the Puren shows evidence
for wave-cut benches, mangrove beaches, brackish lagoons, and hints of an
island fauna. About 1.9 mya, a series of mass mudflows abruptly slid down
a nearby volcanic cone to fill in local coastal areas. These lahars (slurries
of wet ash mixed with larger rocks, tree trunks, and the occasional animal
carcass) raised the local environment high enough to support freshwater lakes
and swamps with higher, dryer ground inland. On top of the lahars, the Sangiran
formation is mostly lacustrine clay with heavy organic content. Fossils of
the first-arriving land mammals appear in these swamp deposits: water-loving
species such as elephants, hippos, and certain deer.
As the Sangiran formation continued to build up, the lakes became shallower
and more productive, and the nearby terrestrial landscapes must have been
lush. Homo erectus arrived to such lakeside environments along with
the first large carnivore, Panthera. As the early humans settled in,
the Sunda coast kept emerging, and the local landscape kept drying.
In yet another abrupt transition about 1.5 mya, local uplift and volcanic
eruption took over the creation of
area landforms.
Local sedimentation changed from lacustrine to fully fluvial or riverine.
The current dome area quickly took on much of its present aspect, a low
terrestrial plateau framed with large volcanoes. This fluvial sediment mantle
is known locally as the Bapang formation.
Active volcanism alternated with more peaceful times during the period
represented by the Bapang formation. During the more energetic episodes,
uplift increased stream gradients, and eruptions charged stream courses with
volcanic detritus. As this sediment was transported seaward, the Sangiran
locale found itself ever more distant from the coast. Thick layers of sand
and gravel in fast-shifting stream beds typify the active phases. In quieter
times, meandering rivers accumulated beds of fine tuffaceous sediment.
We count five two-phased cycles of sedimentation in the Bapang formation.
Most large mammal fossils are found in the coarse sedimentary phases (fig.7).
We also find that the sedimentary cycles lose intensity through time. Rich
bone beds lie just above the Bapang/ Sangiran formations transition zone.
The lower Bapang human fossils here are highly fragmented teeth and jaws.
As one moves up through the Bapang sediment column, rock particles get finer
and the fossils become more complete and well preserved (fig.8)
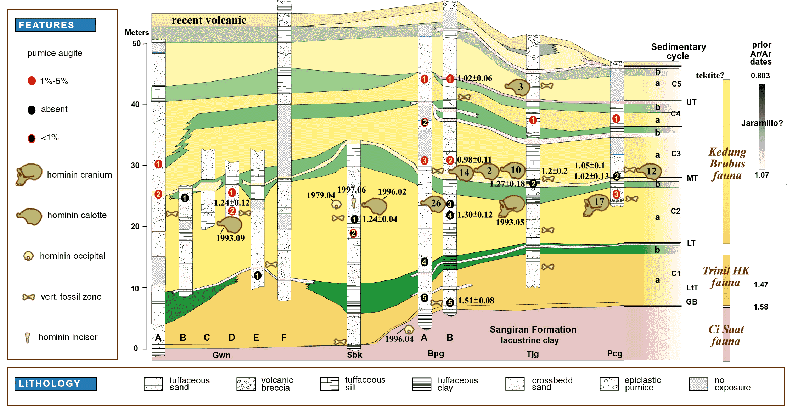
Fig.7: Correlation of Sangiran stratigraphy with hominid finds. Columns
indicate sampled localities, and numbered ovals indicate pumice samples tested
for augite content, hornblende color, and 40Ar/39Ar age analysis. Skulls
found into the mid-1970s are labeled with their “Sangiran” number
(e.g., Sangiran 17). Later finds are referred to by the locality, year, and
month of their discovery (e.g., Tjg 1993.05). Locality abbreviations, left
to right (north to south, with no scale): Gwn - Grogolwetan; Sbk - Sendangbusik;
Bpg - Bapang; Tjg - Tanjung; Pcg - Pucung (after Larick et al. 2001). While the formation names change, Bapang-like sedimentary cycles have continued
right up to the present in Central Java. The human fossils, however, and
most of the accompanying large mammal remains, are not present in the upper
part of the Bapang formation itself. The time of disappearance must be about
800,000 years ago. For reasons not yet entirely clear, the well-entrenched
early human population disappeared from the Sunda south coast after having
lived there for more than a half million years.
In sum, the earliest Homo erectus groups arrived to the coastal swamps
of south-central Sunda between 1.8 and 1.6 mya. At this point the hominids
probably lived slightly inland of the Sangiran locale; their bones washing
downstream after death. About 1.5 mya, fast-flowing streams began building
and cutting beds of coarser sediment. The lower and middle Bapang formation
stream banks represent the landscapes on which Homo erectus actually
lived. About 800,000 years ago, the hominids and most other contemporary
large mammals seem to have left the area. In the meantime, volcanic debris
has continued to accumulate up to the present era.
Age of Sunda Homo erectus: During the last century, age estimates
for the earliest human fossils on Java have ranged from as little as a half
million years to as much as 2 million years. As in other areas of human
evolutionary research, the initial age estimates were young; newer geological
findings and dating methods tend to push back the ages. Our collaboration
with Indonesian geologists and American geochronologists applies the most
advanced radiometric dating methods (Larick et al. 2001). There is now little
question that early Sunda humans arrived toward the end of the Olduvai subchron.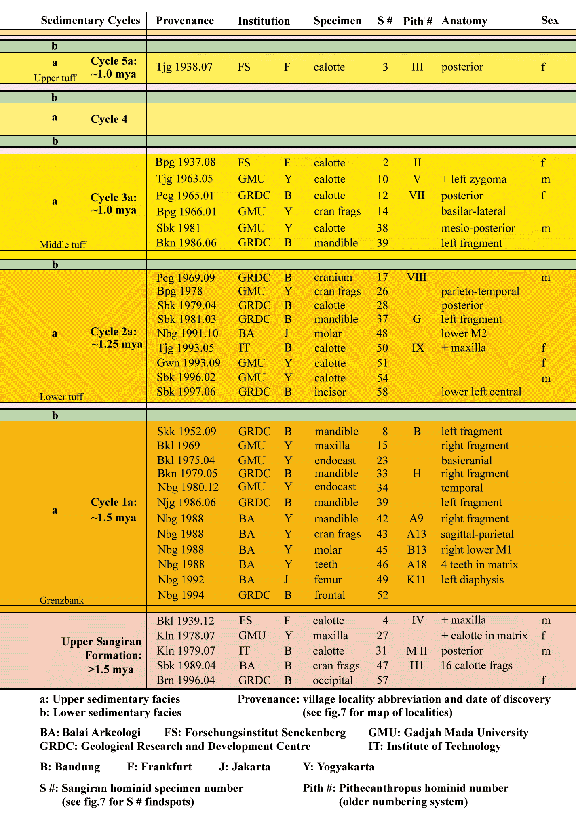
Fig.8: The five two-phased sedimentary cycles of the Bapang Formation
have produced a differential preservation of fossil hominids. The lowest
cycle (1a) with more energy and courser rock particles has yielded fragmentary
teeth and jaws. The upper cycles (2a to 5a) with lower energy levels and
finer sediments have yielded better preserved calottes and crania. This figure
links Homo erectus specimens with the sedimentary cycles that produced
them. The bottom-most level (in tan) in this figure depicts the oldest hominids
found in the Sangiran Dome from the lacustrine Sangiran Formation (after
R. Uribe, R.L. Ciochon, and R. Larick) . The volcanic debris that preserves the Sangiran fossils also provides a means
to understand their age. Small quantities of deep earth minerals, such as
hornblende and plagioclase, are expelled molten during eruptions. At the
moment the droplets cool and crystallize outside the volcano, their potassium
isotopes are maximally unstable. As the phenocrysts become part of the sediment
column, the unstable isotopes decay at a constant rate. Radioactivity decreases
as the stable decay byproducts increase. Given routine assumptions, the relative
proportions of unstable and stable isotopes mark the time since the volcano
has erupted. Geochronologists speak of this ratio as the mineral’s
“eruption age.” If a datable phenocryst and a human fossil have
contemporaneous sedimentary origins, the mineral eruption age approximates
the age of the fossil itself.
But there are some problems. Individual eruptions often produce
“tuffs,” layers of air-fall volcanic ash that provide distinct
horizons within a sedimentary sequence. In East Africa, such tuffs have been
a heavenly gift for geochronologists. Phenocrysts dated from a tuff overlying
fossil beds give a minimum age for any given fossil. Tuff minerals underlying
a fossil bed produce maximum ages. In the humid environments of Southeast
Asia, however, tuff layers are almost always reworked by surface runoff and
stream action. Ash from a number of tuffs may be mixed together and then
deposited against a stream bank from which older fossils are eroding.
These same
conditions also increase the chemical weathering of the volcanic minerals.
In the Sangiran dome, individual crystals are often too weathered to give
true eruption ages. Moreover, Central Javan mineral crystals are often too
small to analyze individually.
In surmounting these problems, we have turned toward clastic (lumps of) pumice,
the sponge-like glass debris common to island arc volcanoes in Southeast
Asia. Pumice clasts usually contain numerous hornblende phenocrysts, and
the encapsulation serves to hold back the weathering process. Moreover, pumice
clasts are often found in the same coarse sediment as the human fossils
themselves. Consequently, our geochronology uses “bulk samples”
of pumice-encapsulated hornblende phenocrysts.
In order to understand the associations of pumice clasts and human fossils,
our Indonesian-American team has undertaken detailed study of the sedimentary
framework for the Sangiran dome (fig.7). We are studying the sedimentary
dynamics of fossil bones and pumice clasts in stream environments and are
analyzing the petrographic variety in volcanic minerals throughout the dome.
The final step is to calibrate these findings with hornblende eruption ages
at a number of stratigraphic levels.
The rich bone
beds of the Bapang formation provide abundant information. In the lowest
Bapang sediments, coarse gravel holds highly fragmented human fossils. The
pumice clasts contain green hornblende that yields eruption ages between
1.51 to 1.47 mya. Above the base, a second cycle of Bapang deposits holds
less fragmented human cranial elements (fig.11). Middle range pumice clasts
have green and brown hornblende crystals with ages from 1.33 to 1.24 mya.
Higher Bapang beds hold the youngest cranium in association with brown
hornblende. The eruption ages cluster around 1 mya.
As already mentioned, the lahars at the base of the Sangiran formation contain
pumice clasts and their hornblende gives eruption ages of 2 to 1.8 mya.
Unfortunately, the Sangiran formation above the lahars has no pumice. For
the present we are not able to include the most important early human fossil
beds directly in our scheme. Nevertheless, the bracketing dated material
indicates that Homo erectus arrived after 1.8 mya and before 1.6 mya.
Our results give the first radiometrically calibrated scheme for the emergence
of this part Sunda, as well as for the arrival, entrenchment, and disappearance
of Homo erectus. This human ancestor occupied south Sunda for at least
a half million years beginning more than 1.6 mya. With an occupation of this
duration, we may speak of an evolutionary sequence for Sunda Homo
erectus. When the large number of Sangiran dome fossils are ordered by
time, it is clear that the Sunda population felt environmental pressures
akin to those of its western Tethys cousins (box 3).
Sunda Homo erectus in Global Context: Fossils representing very early
Homo erectus populations are now known from the highland Rift Valley
of East Africa, the Caucasus Mountains that mediate southeast Europe and
southwest Asia, and from the intensely volcanic slopes of the Sunda subduction
zone. Circum-Mediterranean archaeological sites representing these groups
may be present in northern Algeria (Ain Hanech), Andalusian Spain (Orce),
and the Negev (Erq el Amar). Late Olduvai subchron archaeological sites are
also known on the Himalayan fore slope (Riwat, Pakistan), and in southern
China (Longgupo). The Plio-Pleistocene carnivores associated with humans
are also known from Greece (Mygdonia Basin).
The commonalties among these sites call for a new interpretation of early
Homo erectus. All these sites fall into the transcontinental Tethys
geotectonic corridor, the grand suture at the southern margin of the Eurasian
continental plate with southward extensions into the East African Rift and
the Sunda subduction zone. A global time marker immediately precedes and
overlaps with all sites, the Olduvai subchron (1.96 to 1.79 mya). With the
corridor and the subchron, we can begin to talk about Homo erectus
biogeography as neither African nor East Asian, but as Plio-Pleistocene
Tethys.
The corridor’s linear geotectonic structure includes convergent plate
margins from Iberia to Sunda; rift valleys and plate imbrication zones from
equatorial East Africa to the Caucasus. During the Olduvai subchron, major
Tethys geotectonic events (including the Aullan sea regression in the west
and the emergence of Sunda in the east) served to open virgin territory from
west to east. With large body size, striding gait, carnivorous diet, and
elemental technology - and a relatively small brain - early Homo
apparently found advantage in the realm’s linear structure and the
subchron’s geotectonic instability. Arising in the equatorial western
realm near the beginning of the Olduvai subchron, early Homo erectus
dispersed throughout the realm by or just after subchron end.
Biogeographically, early Homo is a Plio-Pleistocene Tethys lineage,
with long-lived branches in East Africa and East Asia, and possibly in extreme
southern Europe. Morphological variation across the realm is still incompletely
known and potentially very complex. The new Sangiran data indicate that the
two known equatorial populations (Sunda and East African Rift Valley)
encephalized in parallel already by the early Pleistocene. Whatever their
importance for later developments in Homo, neither a large brain nor
a complex stone technology served this hominid’s initial intercontinental
dispersals. Alternatively, the Tethys geotectonic landscapes may have provided
this early human species its greatest advantage and provided the distinction
between it and the numerous preceding australopiths.
References: Djubiantono,
T. and F. Sémah. 1993. “L’île de Java et son peuplement.” In F. Sémah,
A.-M. Sémah, and T. Djubiantono (eds.), Le Pithecanthrope de Java, pp.
12-19, Les Dossiers d’ Archeologie, no. 184. Gabunia L., S.C. Antón, D. Lordkipandze, A. Vekua, A. Justus, and C.C. Swisher III. 2001. “Dmanisi and dispersal.” Evolutionary Anthropology 10: 158-170. Gibert,
J., L. Gibert, C. Ferrández-Canyadell, A. Iglesias, and F. González.
2001. “Venta Micena, Barranco León-5 and Fuentenueva-3: Three
Archaeological Sites in the Early Pleistocene deposits of Orce,
South-East Spain.” In S. Milliken and J. Cook (eds.), A Very Remote Period Indeed: Papers on the Paleolithic Presented to Derek Roe, Oxbow Books, Oxford, UK, pp. 144-152. Huffman, O.F. 2001. “Geologic context and age of the Perning/Mojokerto Homo erectus, East Java.” Journal of Human Evolution 40: 353-362. Koufus, G.D. 1992. “The Pleistocene carnivores of the Mygdonia Basin (Macedonia, Greece).” Annales de Paléontologie 78: 205-257. Larick, R., and R.L. Ciochon. 1996. “The African emergence and early Asian dispersals of the genus Homo.” American Scientist 84: 538-552. Larick, R., R.L. Ciochon, and Y. Zaim. 1999. “Fossil Farming in Java.” Natural History 108(6): 54-57. Larick,
R., R.L. Ciochon, Y. Zaim, Sudijono, Suminto, Y. Rizal, and F. Aziz.
2000. “Lithostratigraphic context for Kln-1993.05-SNJ, a fossil
colobine maxilla from Jokotingkir, Sangiran Dome.” International Journal of Primatology 21: 731-759. Larick,
R., R.L. Ciochon, Y. Zaim, Sudijono, Suminto, Y. Rizal, F. Aziz, M.
Reagan, and M. Heizler. 2001. “Early Pleistocene 40Ar/39Ar ages for
Bapang Formation hominins, Central Jawa, Indonesia.” Proceedings of the National Academy of Sciences, USA 98(9): 4866-4871. Rendell,
H.M., E. Hailwood, and R.W. Dennell. 1987. “Paleomagnetic dating of a
two-million-year-old artefact-bearing horizon at Riwat, northern
Pakistan.” Earth and Planetary Sciences Letters 85: 488-496. Sémah, F., A.-M. Sémah, T. Djubiantono, and H.T. Simanjuntak. 1992. “Did they also make stone tools?” Journal of Human Evolution 23: 439-446. Swisher
III, C.C., G.H. Curtis, T. Jacob, A.G. Getty, A. Suprijo, and
Widiasmoro. 1994. “Age of the earliest known hominids in Java,
Indonesia.” Science 263: 1118-1121 Vekua,
A., D. Lordkipanze, G.P. Rightmire, J. Agusti, R. Ferring, G.
Maisuradze, A. Mouskhelishvili, M. Noiradze, M. Ponce de Leon, M.
Tappen, M. Tvalchrelidze, and C. Zollikofer. 2002. “A new skull of
early Homo from Dmanisi, Georgia.” Science 297: 85-89. Wanpo,
H., R. Ciochon, G. Yumin, R. Larick, F. Qiren, H. Schwarcz, C. Yonge,
J. de Vos, and W. Rink. 1995. “Early Homo and associated artefacts from
Asia.” Nature 378: 275-278. Zhu,
R.X., K.A. Hoffman, R. Potts, C.L. Deng, Y.X. Pan, B. Guo, C.D. Shi,
Z.T. Guo, B.Y. Yuan, Y.M. Hou, and W.W. Huang. 2001. “Earliest presence
of humans in Northeast Asia.” Nature 413: 413-417.
This article appears on pages in Vol.4, No.1 of Athena
Review.
.
|
|