free
trial issue
subscribe
back
issues
Athena Review Vol.2, no.1
Early plant domestication in
Mesoamerica
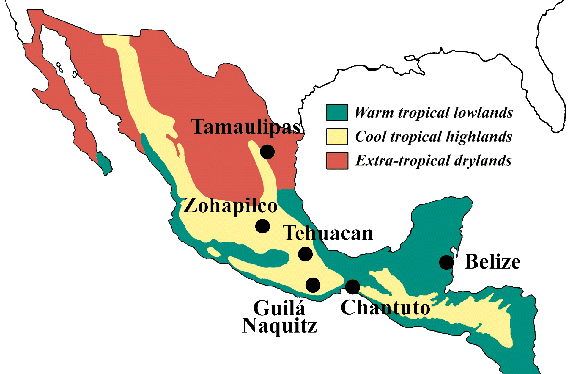 Mesoamerica
(fig.1) is a cultural super-region extending from northern Mexico to Central
America, whose diverse environmental zones (arid; mountainous; tropical lowlands)
each became ancestral areas for major crops ranging from squash, corn, and
beans to cotton and chocolate. Regionally distinct types of food production
emerged as the subsistence base of a wide range of early civilizations, and
the direct precursors of some of today's most important crop plants. Much
of their origin, however, still remains a mystery.
Mesoamerica
(fig.1) is a cultural super-region extending from northern Mexico to Central
America, whose diverse environmental zones (arid; mountainous; tropical lowlands)
each became ancestral areas for major crops ranging from squash, corn, and
beans to cotton and chocolate. Regionally distinct types of food production
emerged as the subsistence base of a wide range of early civilizations, and
the direct precursors of some of today's most important crop plants. Much
of their origin, however, still remains a mystery.
[Fig.1: Environmental zones and major sites of early plant cultivation
in Mesoamerica (after McClung 1992 and MacNeish 1992).]
Domestication versus Cultivation: A domesticated plant, strictly defined,
is one whose reproductive success depends on human intervention. It is often
quite difficult to apply this term accurately to plant taxa represented in
the archaeological record. Archaeobotanical specimens (ie., charred cobs
of corn or maize) are recognized as possible early cultigens because they
possess attributes that fall in between those of known wild progenitors and
those of modern domesticates. In some instances, anatomical features may
be present that suggest that the species would do poorly on its own in the
wild, and, therefore, are unlikely to have been the result of natural selection.
For example, inefficient seed dispersal mechanisms (as in the maize ear),
while greatly diminishing a plant's chances for successful reproduction in
the wild, are always advantageous for seed-harvesting humans, due to more
retention of seeds in the pod. In a great many instances, however, the
deteriorated and fragmentary plant remains recovered from archaeological
contexts lack the definitive anatomical clues allowing determination of just
how far along the road to "true" domestication the specimen may have been.
Wild-Food Production: In contrast to collection (also called foraging
or gathering), wild-food production means behavior intended to alter the
abundance and distribution of plant resources. The term, while somewhat
misleading if we take "production" in the conventional sense of increasing
yield, remains a useful concept for energy investment in plant resource
extraction, involving a group of strategies designed to reduce collection
time. This is mainly done by reducing travelling time to the source areas
in various ways. One is the protection of known stands of exploitable plants
by "tending," to assure a continous availability at known, convenient locales.
Other intensive gathering or "tending" methods include weeding, discouraging
predators, pot-irrigation, and limited harvesting to ensure reproduction.
Another obvious time-saving solution and precursor to domestication is the
direct transplantation of desirable resources closer to a camp or settlement.
Probable candidates for transplantation are mainly small trees or bushy plants
with a perennial growth habit, whose produce (sap, fiber and wood as well
as edible portions) is readily harvested and processed for consumption. In
Mesoamerica this fits such plants as avocado, cacao, capulín, ciruela,
cotton, mesquite, papaya, ramón, zapote blanco, and zapote negro,
among others. Other easily transported, edible species include cacti (ie.,
prickly pear), maguey, and fruit-bearing palms such as coyol. Many, though
not all, of these species can be propagated with cuttings.
To document early transplantation, it must be demonstrated that a species
occurs in prehistoric sites outside of its natural habitat. Good evidence
for this practice exists from some occupation strata in the dry cave sites
of the Tehuacán Valley .from at least 3500 BC and perhaps much earlier
(with dates for wild avocado proposed for ca.7000-6500 BC). Archaeologists
agree that the remains of avocado, ciruela, coyol, zapote blanco and zapote
negro represent exotics introduced by humans into the Tehuacán Valley.
All of these species are native to considerably more mesic (wetter) environments,
and their establishment in the semiarid Tehuacán area would have required
some irrigation (McClung 1992; Pearsall 1995; C. Smith 1987).
The Tehuacán cave deposits are extraordinary for their preservation,
richness and depth. More usual is that the practices of tending and
transplantation may be archaeologically undetectable. Transplantation does
not necessarily involve the crossing of biotic zones. More often, traces
of plants we suppose to have been exploited regularly by prehistoric peoples
simply do not turn up in the excavated material for one reason or another.
The ramón seed, for example, a ubiquitous and valuable resource available
to occupants of the tropical lowland forests, generally is not found in
archaeological deposits, and is not likely to be found owing to its soft,
highly perishable seed coat (Fedick 1995). Thus, we are unable to track its
history as a cultigen. The archaeobotanical record for Mesoamerica is full
of such huge gaps regarding when and how the dispersal of economically useful
plants were effected via human cultural practices.
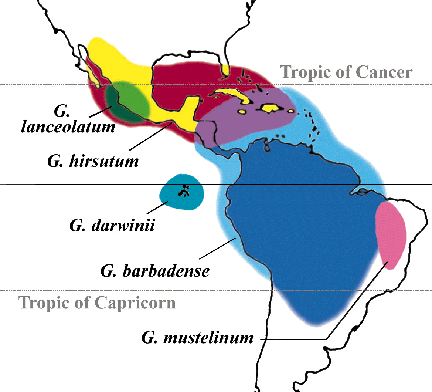 Cotton and
its dispersal: A prime example of this incomplete state of knowledge
on plant origins is cotton (Gossypium spp.), which has both Old and
New World species. We do not know when or where prehistoric Mesoamericans
first began to appreciate the utility of the cotton plant, even though something
is known about the complex distribution of wild populations in the New World.
As far as has been determined, wild cotton occurs in distinctive and
restricted littoral (shoreline) or related habitats. It must have taken
considerable time for the species to have dispersed so widely throughout
tropical and subtropical Mesoamerica, and to have developed the long-fibered
seeds for which it is so highly valued. The seeds of the littoral wild cottons,
perennials which do not propagate vegetatively, have very sparse fibers.
The oldest known archaeological samples of Mesoamerican cotton are from the
Tehuacán Valley, dated 3500-2300 BC; these represent a fully domesticated
form, Gossypium hirsutum (Smith and Stephens 1971).
Cotton and
its dispersal: A prime example of this incomplete state of knowledge
on plant origins is cotton (Gossypium spp.), which has both Old and
New World species. We do not know when or where prehistoric Mesoamericans
first began to appreciate the utility of the cotton plant, even though something
is known about the complex distribution of wild populations in the New World.
As far as has been determined, wild cotton occurs in distinctive and
restricted littoral (shoreline) or related habitats. It must have taken
considerable time for the species to have dispersed so widely throughout
tropical and subtropical Mesoamerica, and to have developed the long-fibered
seeds for which it is so highly valued. The seeds of the littoral wild cottons,
perennials which do not propagate vegetatively, have very sparse fibers.
The oldest known archaeological samples of Mesoamerican cotton are from the
Tehuacán Valley, dated 3500-2300 BC; these represent a fully domesticated
form, Gossypium hirsutum (Smith and Stephens 1971).
[Fig.2: Map of New World cotton species (after Fryxell 1979).]
Cotton may possibly have been introduced to Mesoamerica as an already
domesticated form from the southern hemisphere where, in coastal Ecuador
and Peru, cultivated cotton of another species (G. barbadense) has
been documented for much earlier periods. Yet most views favor the independent
domestication of G. hirsutum in Mesoamerica (Fryxell 1978; Heiser
1990; Pickersgill 1989). Confirmation of this judgment is needed from the
archaeological record.
Adding to the general uncertainty concerning the earliest attempts to manage
wild-food production is that mere collection behaviors may have side effects
which contribute to the domestication process. Transportation of seed-bearing
fruits to a settlement may enable a species to spread beyond its original
"natural" range. Newly established populations may develop characteristics
different from those of parents if environmental conditions (ie., altitude,
soil conditions, precipitation) foster such adaptations, or if "unconscious
selection" operates because collectors habitually harvest specimens with
particular traits such as large size, ease of detachability from the stem,
or lack of spines. The end result is to promote speciation, even when no
food production techniques were practiced.
Straightforward interpretations of archaeobotanical assemblages are rarely
possible. Social factors including group size, division of labor, and degree
of sedentism, as well as technological factors, all must be considered in
reconstructions of subsistence related behaviors. Wild-food production could
fit within most social organizations, but presumes at least a single season's
residency at one place. Fully mobile groups obtain no subsistence advantage
by investing time in propagating or maintaining resource stands which are
visited infrequently. The Coxcatlán Cave data from the Tehuacán
Valley show a predominance of fruits gathered from perennials (trees, agave,
and cacti) in the earlier levels of ca. 7000-3500 BC (MacNeish 1992; Pearsall
1995). Wild grasses and annuals were certainly available but appear not to
have been high priority foodstuffs. Likewise, no grasses at all were reported
for the Archaic period (8900-6670 BC) occupations at Guilá Naquitz
Cave in the Oaxaca Valley near the town of Mitla (C. E. Smith, 1986). Kent
Flannery (1986) and colleagues, who excavated Guilá Naquitz, emphasize
the selectivity of the occupants with respect to number of plants exploited
compared with the great variety available. Archaeologist Brian Hayden (1995)
warns that, since sites like Coxcatlán Cave and Guilá Naquitz
"were peripheral to the main valley ecosystem[s] and may have been visited
only infrequently," the botanical remains recovered may not represent the
full range of resources utilized. This idea warrants serious consideration.
Nonetheless, there is no reason to believe that, in a moderately bountiful
environment, people would have had no food preferences and simply consumed
indiscriminantly whatever lay in their path. The choice of easily harvested,
relatively large food packages that required minimal processing prior to
consumption fits behavioral expectations for band-level foraging groups with
a fairly high degree of residential mobility.
Cultivation: The line drawn between wild-food production and cultivation
can be a thin one that rests mostly on assumptions regarding the effort made
by prehistoric peoples to manipulate the growth of plants. According to Harris,
cultivation, as the second level of energy investment, involves systematic
soil preparation and planting as well as tending and harvesting. Initially,
cultivation can be thought of as an extension of the collection-time reduction
principle, where the major goal is ensuring availability. However, as more
of the time saved by not collecting is rerouted into field and crop management,
it will be seen that such attention often improves the health and yield of
the subject plants. Cultivation, therefore, ultimately comes to represent
a commitment to improving the product itself and its productivity, in addition
to availability.
Depending on the genetic malleability of the species, selection for desirable
traits will become a conscious and prominent aspect of the process. In the
archaeological record, the plant taxa identified as the earliest cultigens
are not the same sort of plants that were the focus of wild-food production.
In general, they are annuals or short-lived perennials that readily colonize
disturbed habitats, such as human occupation sites. It also happens that
most of the early Mesoamerican cultigens represent much smaller food packages
than, for example, collected tree fruits, and require extra time to process
for consumption. The energy trade-off only makes sense where there is at
least part-time sedentism and a group sufficiently large for some members
to devote themselves to time-consuming tasks of processing small food items.
Cultivated Plants: Several species of the squash family (Cucurbitaceae;
genus Cucurbita) are among the earliest known cultigens. Cucurbita
pepo, the pumpkin squash (fig.5), has been dated to 8000-7000 BC at
Guilá Naquitz and at Ocampo Cave in Tamaulipas to about 7000-6500
BC (Heiser 1989; B. Smith 1997). The specimens were identified as cultivated
forms on the basis of morphological criteria and, as the Guilá Naquitz
material has been dated directly, C. pepo is confirmed as the oldest
known cultigen for Mesoamerica (see AR, I,3). C. pepo, along with
the other domesticated cucurbits known from archaeological contexts (C.
argyrosperma, C. ficifolia, C. lundelliana and C.
moschata), appear to have arisen from different wild species. Although
none of the wild cucurbit progenitors has been identified with certainty,
it  is believed
that the centers of cultivated species diversity are the areas of origin.
The ancestral squashes would have been cultivated principally for their
nutritious edible seeds, since their scant bitter flesh would have required
repeated boilings to eliminate the unpalatable cucurbitacins (Nee 1990).
Selection for larger, fleshier, non-bitter fruits could have been accomplished
relatively rapidly given the growth habit of these species and their capability
of self-pollination. Another member of the Cucurbitaceae family, Lagenaria
siceraria, the calabash or bottle gourd, is thought to have been brought
under cultivation for its usefulness as a container, owing to its unusually
thick rind, rather than for food. The bottle gourd also occurs very early
in several Mesoamerican archaeological deposits (Tamaulipas ca. 7000 BC,
Tehuacán ca. 5050 BC, Guilá Naquitz ca. 7000 BC). It is usually
encountered as rind fragments and it is not known exactly when
Lagenaria became domesticated.
is believed
that the centers of cultivated species diversity are the areas of origin.
The ancestral squashes would have been cultivated principally for their
nutritious edible seeds, since their scant bitter flesh would have required
repeated boilings to eliminate the unpalatable cucurbitacins (Nee 1990).
Selection for larger, fleshier, non-bitter fruits could have been accomplished
relatively rapidly given the growth habit of these species and their capability
of self-pollination. Another member of the Cucurbitaceae family, Lagenaria
siceraria, the calabash or bottle gourd, is thought to have been brought
under cultivation for its usefulness as a container, owing to its unusually
thick rind, rather than for food. The bottle gourd also occurs very early
in several Mesoamerican archaeological deposits (Tamaulipas ca. 7000 BC,
Tehuacán ca. 5050 BC, Guilá Naquitz ca. 7000 BC). It is usually
encountered as rind fragments and it is not known exactly when
Lagenaria became domesticated.
[Fig.3: Seed of an early cultivated squash, Cucurbita pepo
(after B. Smith 1997).]
The evidence for bean (Phaseolus spp.; Fabaceae) cultivation is not
of such antiquity as that for squash, although ultimately beans became the
high protein staple of the Mesoamerican diet. It is sensible to presume that
beans were cultivated quite early given that they have many of the same
characteristics that made the squashes such willing domesticates. Wild beans
had a significant presence in the older levels at Guilá Naquitz of
ca. 8900-6500 BC (Kaplan 1986). The kinds of remains suggest that in Preceramic
times, beans had been harvested for their tender shoots or pods. Cultivation
and selection for certain seed characteristics probably followed the introduction
of ceramic technology and attendant cooking techniques that enhanced the
seeds' desirability as food. The transition from noncultivated to cultivated
forms is not clear in the archaeological record, but recent biochemical
investigations strongly indicate that the common bean, Phaseolus
vulgaris, evolved independently in Mesoamerica from the common wild bean,
with a likely center of domestication in Jalisco in West-Central Mexico (Gepts
1990; Singh et al. 1991).
In the tropical lowlands, starch in the diet is more likely to have come
from roots or tubers than from cereals. Unfortunately, the archaeological
record for lowland Mesoamerica has not provided generous amounts of data
with regard to possible root crops. The case for manioc (Manihot
esculenta; Euphorbiaceae) cultivation is the strongest, especially since
some phytogeographers favor a Mexican origin for the species (Hawkes 1989;
C.E. Smith 1987). Variants with large underground storage organs would survive
best in areas with a marked dry season where such reserves would have been
adaptive. But, few sites have yielded direct evidence for manioc consumption.
Root crops are rarely found as carbonized remains and desiccated specimens
are extremely hard to identify. The starchy pulp that constitutes the comestible
portion has hardly any distinctive morphological characteristics. M.
esculenta pollen has been identified in freshwater cores from northern
Belize. On the basis of the pollen data, the researchers estimated a date
of ca. 3400 BC for the introduction of manioc. Since this date roughly coincides
with that for the appearance of maize as well as indications of local
deforestation, they conclude that the manioc was cultivated (Pohl et al.
1996). Manioc has been identified for Formative Period Cuello, also in Belize
(Fedick 1995). These identifications are open to doubt, and the major arguments
for manioc cultivation still rely on inferences based on availability, the
weedy tendencies of the plant, and the technical simplicity of cultivating
it. We do not know whether sweet and/or bitter varieties were consumed, though
the latter demands a great deal more processing prior to consumption. The
claim for manioc at Tehuacán is extremely shaky, as is the claim for
yam (Xanthosoma sp.; Araceae) at Tikal, and no evidence has yet come
to light for the prehistoric cultivation of jícama (Pachyrrhizus
erosus; Leguminoseae).
Many obstacles present themselves to archaeologists wishing to document the
early practice of cultivation, not the least of which is the lack of
straightforward evidence. The most convincing proof of cultivation is, of
course, intentional selection for desired traits. Often, however,
archaeobotanists have only the most limited kinds of morphological indicators
with which to make such determinations, such as an increase in seed size
over time. As so many of the cultigens were initially ruderals (ie, growing
in trash middens), it is very possible that they began to acquire such
characteristics as an increase in fruit size simply because they were doing
so well in an amenable environment. For example, the highly nitrogenous soils
associated with human habitation sites can allow plants to achieve larger
than "normal" size without any intentional selection for large size on the
part of incipient cultivators (ie., incidental domestication).
The case for purposeful cultivation is made more plausible by combining
archaeobotanical and palaeoenvironmental information with an understanding
of the social context in which cultivation behaviors are believed to have
occurred. As noted above, time taken from collection to devote to cultivation
presupposes certain social organizational features that make cultivation
an efficient and attractive strategy. Still, the practice of cultivation
covers a great range of energy input levels, from marginal "back-door" gardening
to a high degree of dependence on the products of cultivation. Many of the
products of cultivation are storeable resources. Evidence of storage, in
the form of food caches or specialized facilities, can help us gauge the
relative importance of cultivation as a subsistence strategy (see also Great
Basin article, this issue).
Agriculture: In Harris' "scale" of people-plant interactions, agriculture
represents the highest level of energy investment. Unlike incipient cultivation,
where hunting and gathering remain important, once the stage of agriculture
is reached, the group is solidly dependent on food production for the greatest
proportion of its subsistence needs. According to Harris, what differentiates
cultivation from agriculture is not precisely the kinds of behavior involved,
but rather the scale at which they are practiced. Agriculture is just highly
intensive cultivation that, on the whole, leaves little time for other
subsistence methods. Its practice presupposes the prior existence of suitable
plants for intensive cultivation, or domesticates. So, whereas the activity
of selection is the primary clue to the beginnings of plant cultivation,
it is the goal of increased productivity of species with known desired qualities
that defines agricultural practice.
For reasons of scale, evidence for agriculture is highly visible in the
archaeological record and is unlikely to be misinterpreted. From the standpoint
of the botanical remains, the inventory of food items will be dominated by
one or a few cultigens. In Mesoamerica, only one domesticated plant, maize
(Zea mays subsp. mays; Gramineae) ever became the object of large-scale,
intensive agriculture. Often readily detectable, as well, are related
modifications to the landscape from clearance, ground levelling via terracing
or other means, and irrigation or other water management schemes (ie., diversion,
drainage, and containment). Perhaps most significant is our knowledge that
sizeable sedentary populations could not be sustained in many environments
without large-scale food production.
Maize and its origins: Despite the ease with which the practice of
agriculture can be identified archaeologically, the transition to agriculture
remains one of the most poorly understood phenomena in Mesoamerican prehistory.
This is largely because the evolutionary history of maize needs unravelling.
For agriculture to be successful, the cultivated crop must possess certain
attributes in order to justify the enormous efforts expended in land tillage,
planting, tending and harvesting. Above all, the yield of usable produce
per plant must be quite high. This criterion is met by maize by virtue of
its remarkable ear and is the reason maize became the most important crop
by far throughout Mesoamerica. The question is, how and when did the fruitful
maize cob develop?
Referring to these mysterious origins, Kent Flannery (1986) has called maize
the "most controversial and most enigmatic of any major cultivated plant."
All specimens of primitive maize known from archaeological sites are domesticated
forms. Maize is not included in the taxa considered above under "wild-food
production," nor does it appear on the list of the earliest cultigens. This
is because it simply wasn't there. Identifying the wild progenitor(s?) of
maize means reconstructing genetic events with little material corroboration,
and so has been the subject of much speculation and debate for at least 100
years. Opinions are numerous, but currently the three leading hypotheses
are: the Wild Maize Hypothesis, the Orthodox Teosinte Theory, and the
Catastrophic Sexual Transmutation Theory (as shown in Benz and Iltis 1992).
The Wild Maize Hypothesis claims that there must have once been a wild race
of maize, now extinct, from which the cultivated form evolved. This theory
explains the development of the cob by asserting that the hypothetical ancestor
was a naturally occurring pod-corn with vertical rows of kernels (polystichy).
The problem with this theory is that no fossilized examples of the putative
ancestral maize has ever been found. Prior to domestication this wild maize
must have been exploited to some extent by prehistoric people and, therefore,
should have found its way into the archaeological record.
 The Orthodox Teosinte
Theory says that maize was derived from the wild grass teosinte (Zea mays
mexicana). In some classifications annual teosinte is treated as a separate
species (Zea mexicana). The basis of this argument is that teosinte
is the closest known relative of maize. Some reject teosinte as the direct
ancestor because the very minute female inflorescences ("ears") of teosinte
seem unlikely to have been transformed into the huge polystichous maize ears
by means of human selection. A variant of the Teosinte Theory, which serves
to reconcile it with the Wild Maize Hypothesis, is that maize is a hybrid
of teosinte and a wild maize ancestor. In other versions teosinte and another
related grass are the hybridizing pair (Mangelsdorf 1986).
The Orthodox Teosinte
Theory says that maize was derived from the wild grass teosinte (Zea mays
mexicana). In some classifications annual teosinte is treated as a separate
species (Zea mexicana). The basis of this argument is that teosinte
is the closest known relative of maize. Some reject teosinte as the direct
ancestor because the very minute female inflorescences ("ears") of teosinte
seem unlikely to have been transformed into the huge polystichous maize ears
by means of human selection. A variant of the Teosinte Theory, which serves
to reconcile it with the Wild Maize Hypothesis, is that maize is a hybrid
of teosinte and a wild maize ancestor. In other versions teosinte and another
related grass are the hybridizing pair (Mangelsdorf 1986).
[Fig.4: Single-rowed ear of Teosinte (after Mangelsdorf 1976).]
Recently, considerable support for teosinte as the direct ancestor of maize
has accumulated as a result of DNA and isozyme (genetically variant enzyme)
studies that provide measures of genetic distance much more reliable than
judgments based on morphological characters. These investigations have revealed
such a high degree of similarity between teosinte and maize with respect
to the presence and frequency of specific molecules that a close phylogenetic
relationship must be a matter of fact (Doebley 1990, 1992). In other species
such a level of similarity might not be so remarkable, but maize is so
exceptionally polymorphic for the traits studied (Hoisington 1992) that the
calculations of genetic distance are very refined. Moreover, the molecular
evidence has identified a particular race of teosinte (Z. mays subsp.
parviglumis), a perennial grass, more similar to maize than any other
race of teosinte. This finding supplies a probable geographic origin for
maize domestication, as well as an immediate ancestor. The wild Z. mays
subsp. parviglumis race that best matches the isozyme profile of maize
has a limited distribution today in the central portion of the Balsas River
drainage in northern Guerrero, eastern Michoacan and western Mexico states
(Doebley 1990). Archaeologists in general are impressed by the statistical
soundness of the perennial teosinte's claim as the progenitor of maize and
the view is widely (though not unanimously) accepted in the discipline. The
Orthodox Teosinte Theory, however, does not resolve the questions about the
development of the maize cob. Botanists are harder to convince and continue
proposing other evolutionary reconstructions.
The Catastrophic Sexual Transmutation Theory, first advanced by Hugh Iltis,
offers an explanation for the development of the maize ear while completely
accommodating the Teosinte Theory. Briefly, the theory holds that through
a series of genetic mutations the staminate teosinte tassel spike (the male
reproductive organ) was transformed into the unisexual distillate maize ear
(the female organ). This contrasts with conventional thinking that has the
maize ear developing from the teosinte ear. But Iltis claims that there is
greater structural homology between the teosinte spike and the maize ear
than between the two ears. Furthermore, the maize ear exhibits vestiges of
its former male anatomy. The radical and rapid anatomical changes associated
with transmutation provides explanations for the late development of maize
as a cultigen and the failure to find "wild maize" in the fossil record.
Such evolutionary adaptations may have been encouraged by volcanic activity
typical of the Balsas River region in the recent past, and sexual transmutation
may have occurred as late as 8000 years ago (Benz and Iltis 1992). The
Catastrophic Sexual Transmutation Theory is a complex argument not uniformly
embraced by maize researchers (Wilkes 1989).
 How the maize
ear developed is a critical question for the timing of maize domestication
but, as of yet, no consensus exists. Numerous examples of primitive maize
obtained from archaeological excavations suggest that cultivated maize diffused
quite swiftly throughout Mesoamerica, especially if we accept a single center
of domestication and a date of about 8000-6000 BC. Tehuacán in the
central highlands of Mexico has yielded the oldest remains of cultivated
maize in Mesoamerica, dated to 5050 BC (calibrated) by accelerator mass
spectrometry dating of actual specimens. Racial diversification arose as
a result of adaptations to different environmental conditions and human selection
for traits related to productivity, taste and cooking properties (DeWet 1992).
Within a few thousand years, maize had become a staple food crop throughout
Mesoamerica, and was a main subsistence base of the Maya, Aztec, and related
early civilizations.
How the maize
ear developed is a critical question for the timing of maize domestication
but, as of yet, no consensus exists. Numerous examples of primitive maize
obtained from archaeological excavations suggest that cultivated maize diffused
quite swiftly throughout Mesoamerica, especially if we accept a single center
of domestication and a date of about 8000-6000 BC. Tehuacán in the
central highlands of Mexico has yielded the oldest remains of cultivated
maize in Mesoamerica, dated to 5050 BC (calibrated) by accelerator mass
spectrometry dating of actual specimens. Racial diversification arose as
a result of adaptations to different environmental conditions and human selection
for traits related to productivity, taste and cooking properties (DeWet 1992).
Within a few thousand years, maize had become a staple food crop throughout
Mesoamerica, and was a main subsistence base of the Maya, Aztec, and related
early civilizations.
[Fig.5: Aztec maize offering depicted in the early 16th century Codex
Borbonicus.]
(Abbreviated from the article by Virginia M. Betz on pp.24-31 in
Athena Review, Vol.2, no.1.)
Athena Review Image
Archive™ |
Paleoanthropology in
the News
| Guide
to Archaeology on the Internet |
Free issue
|
Back issues
Main index of Athena
Review |
Subject Index
| Travel
Pages |
Galleries and
Museums |
Ad
rates |
Current issue
index |
Exhibition Reports
index
Copyright © 1996-2002 Athena
Publications, Inc. (All Rights Reserved).